
Alcohol Dependence Induces CRF Sensitivity in Female Central Amygdala GABA Synapses

 ,
, 
Abstract
:1. Introduction
2. Results
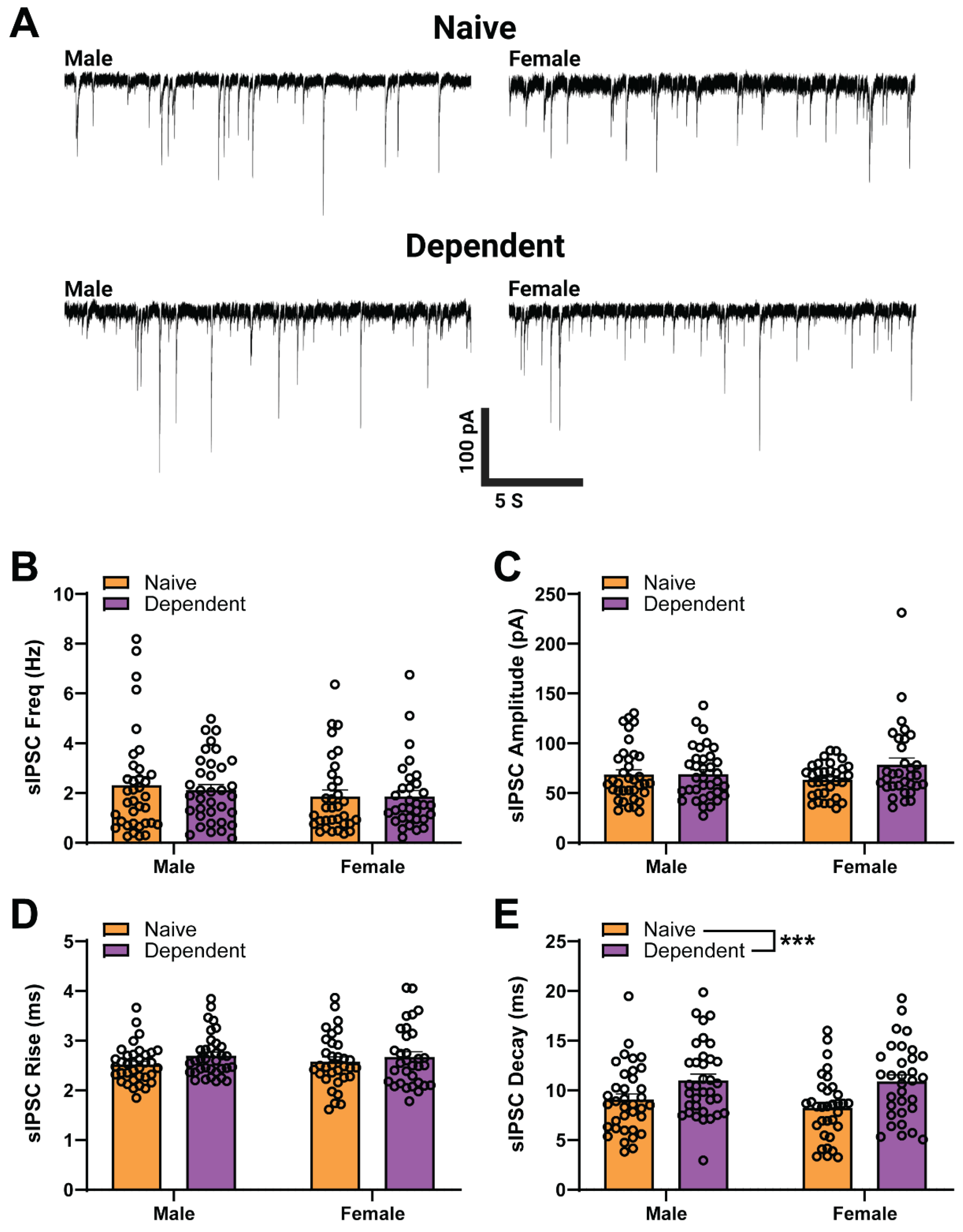
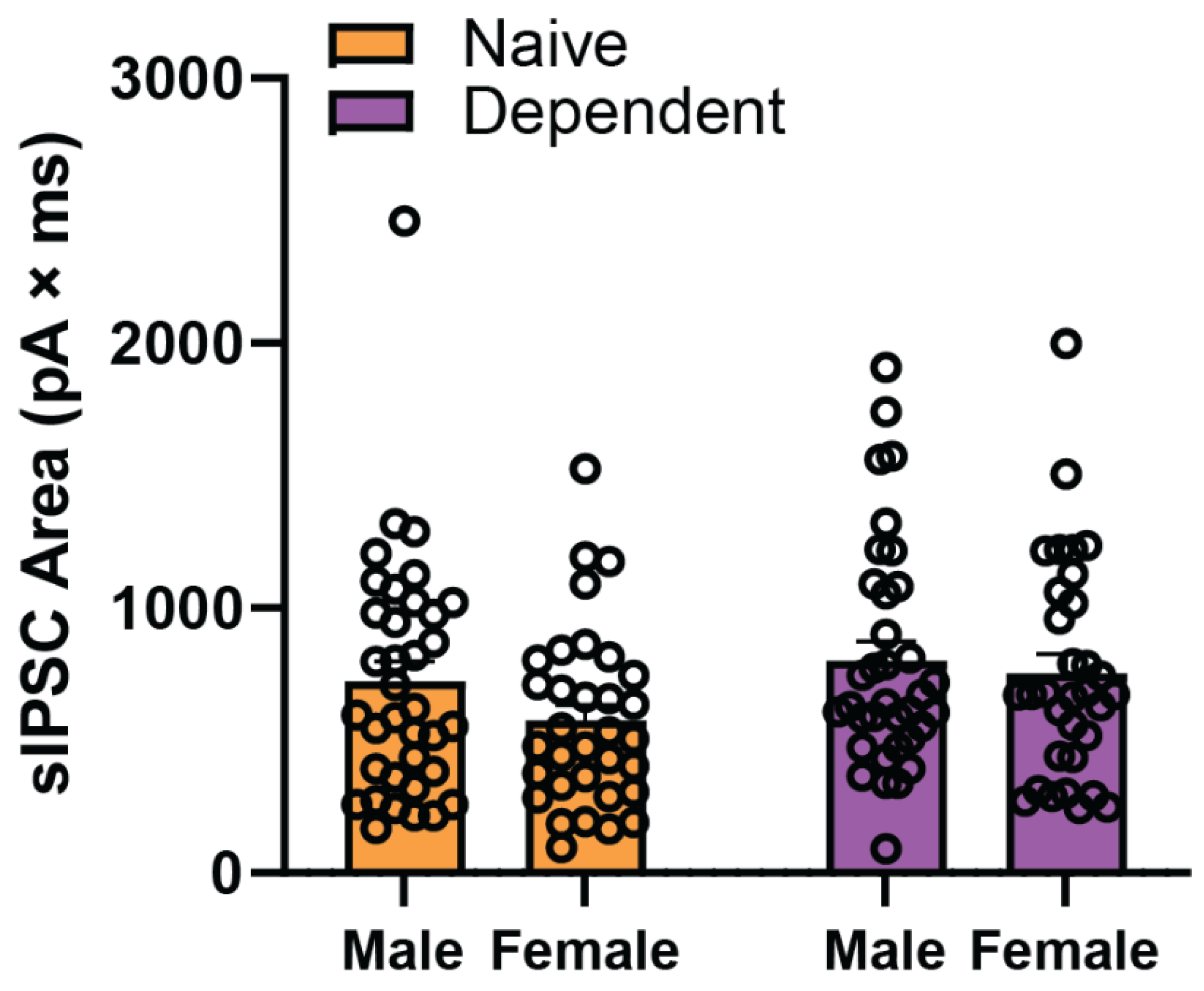
2.1. Sex Differences in Baseline sIPSC Kinetics of CeA GABA Synapses
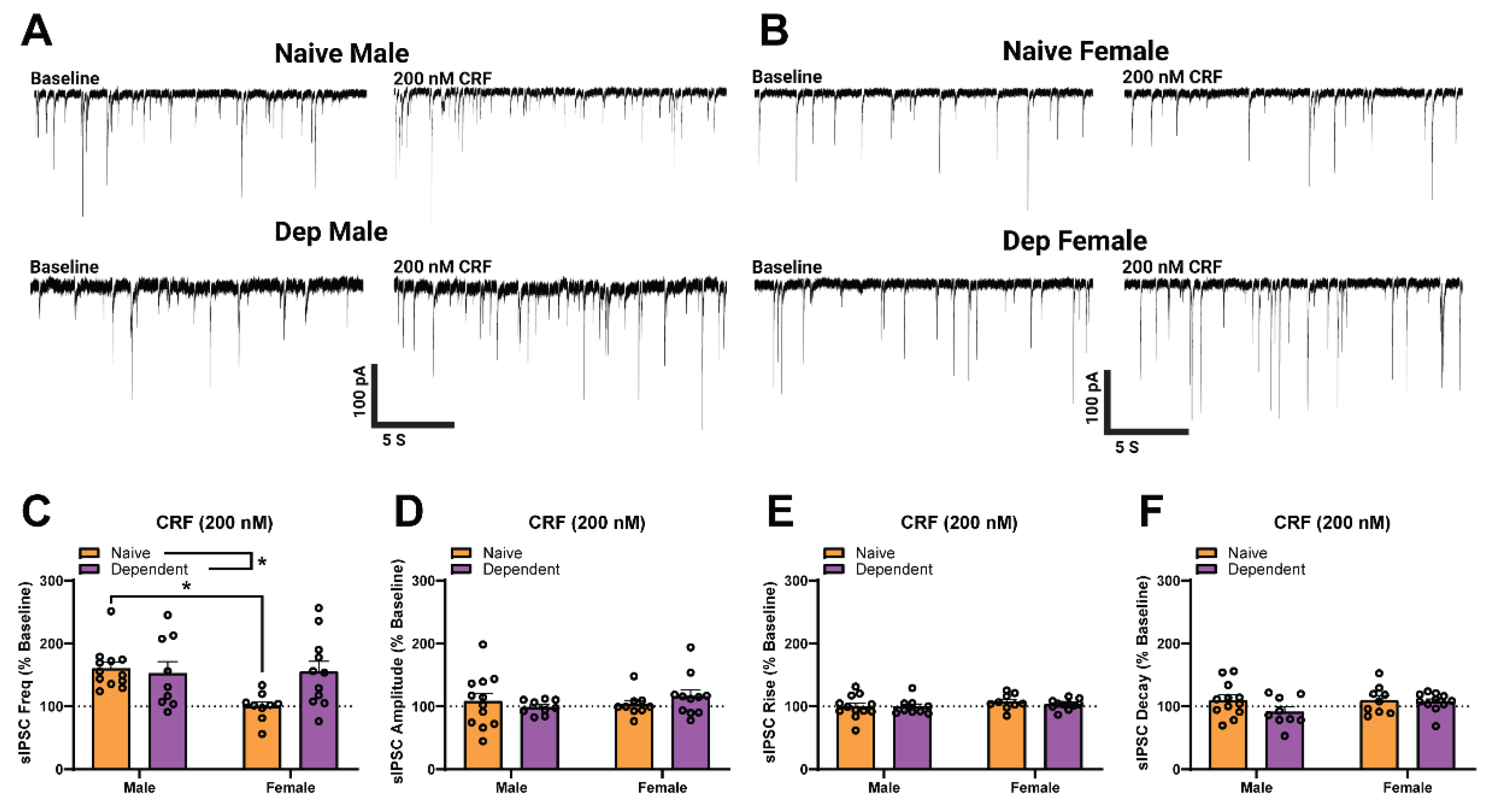
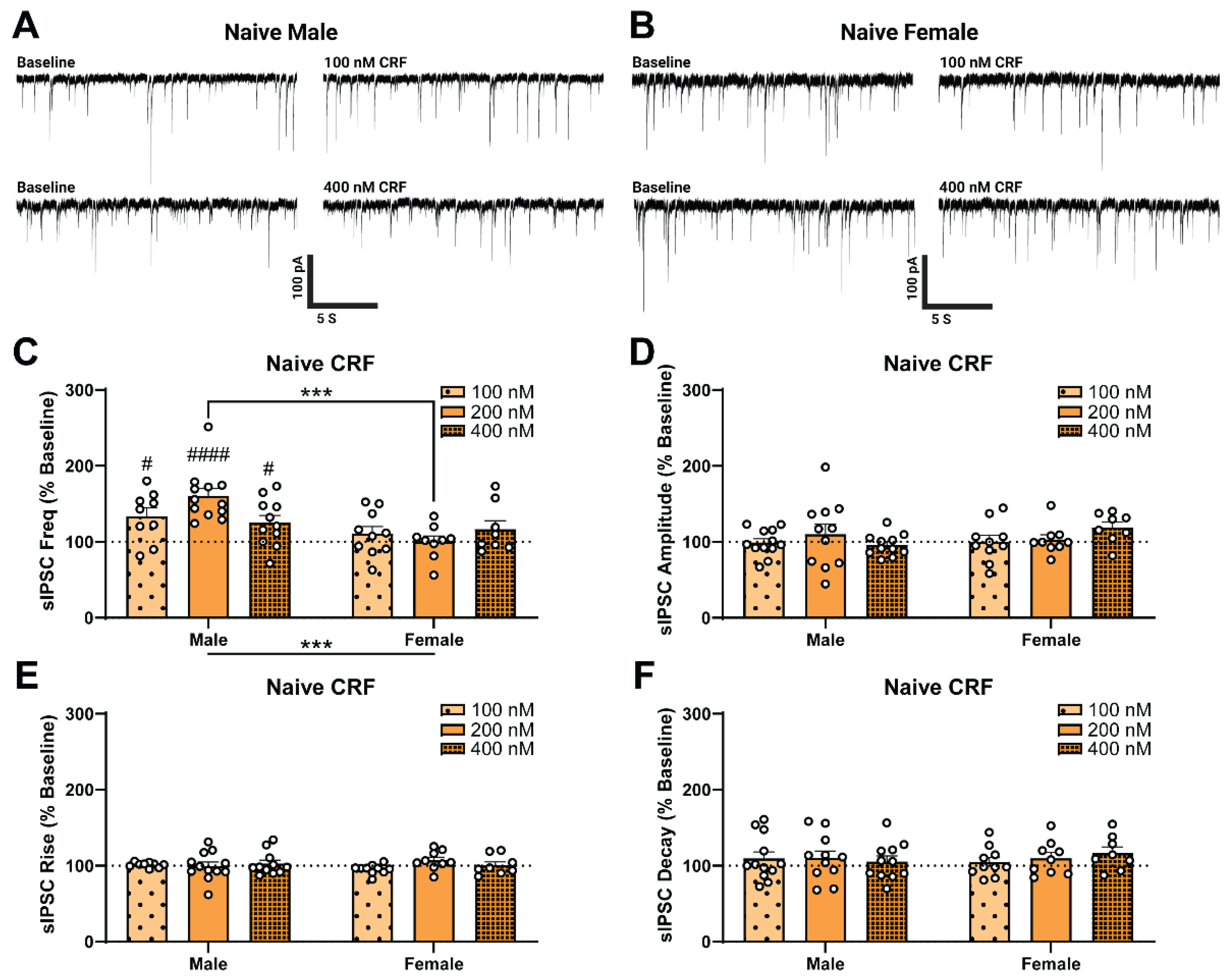
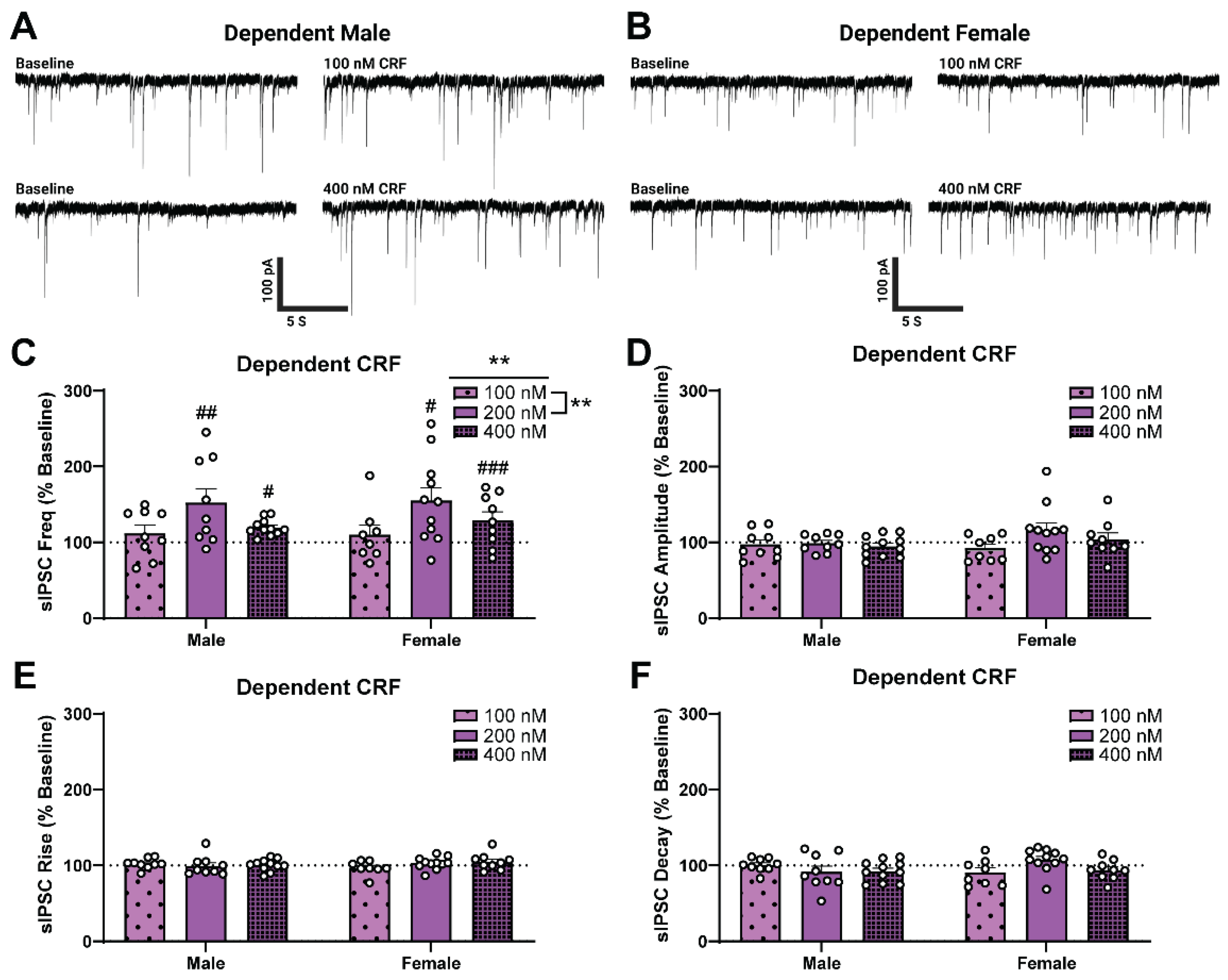
2.2. Alcohol Dependence Induces Responsivity of Female CeA GABAergic Synapses to Acute CRF Application
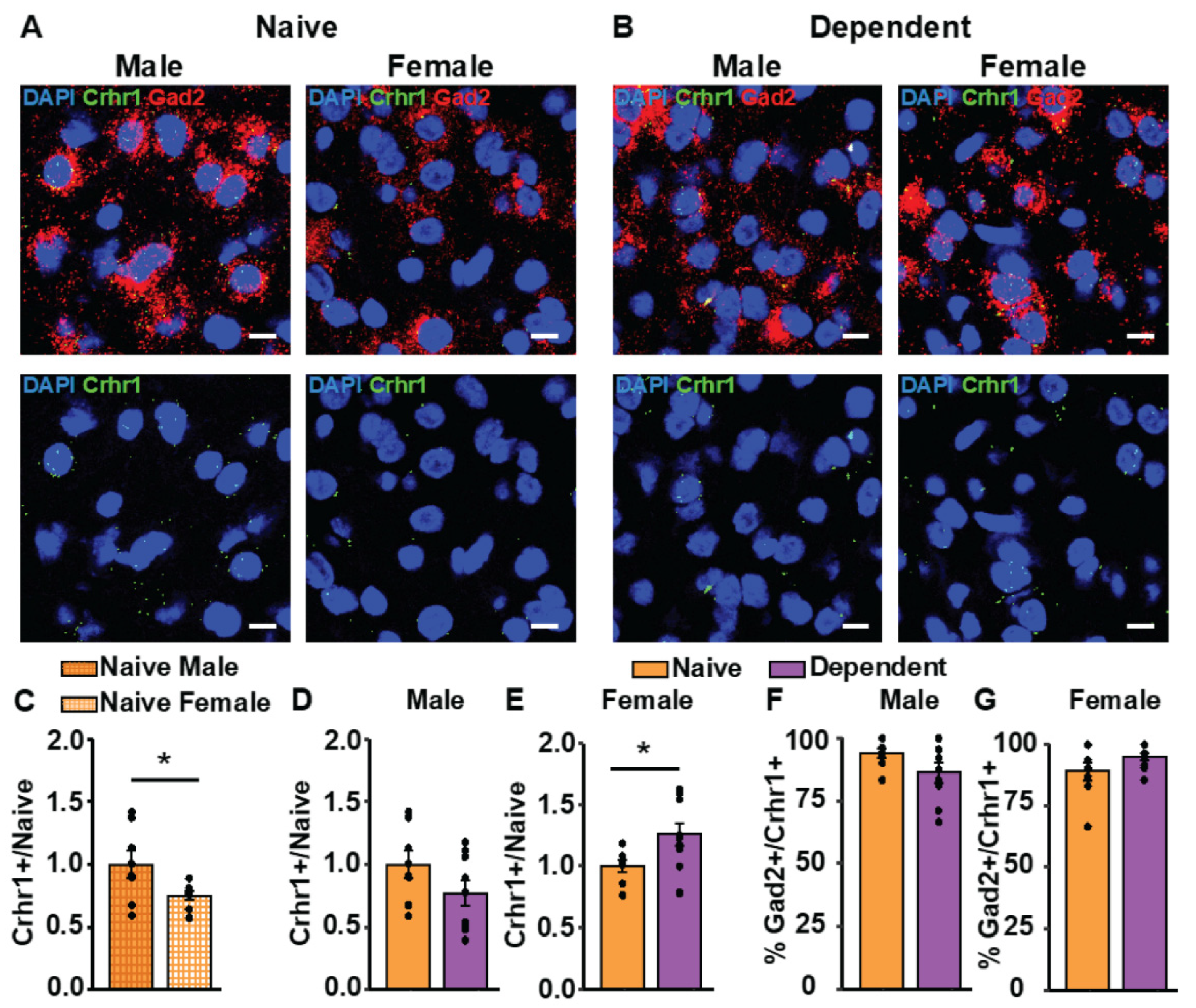
2.3. Alcohol Dependence Alters CRF1 Receptor Expression in Females
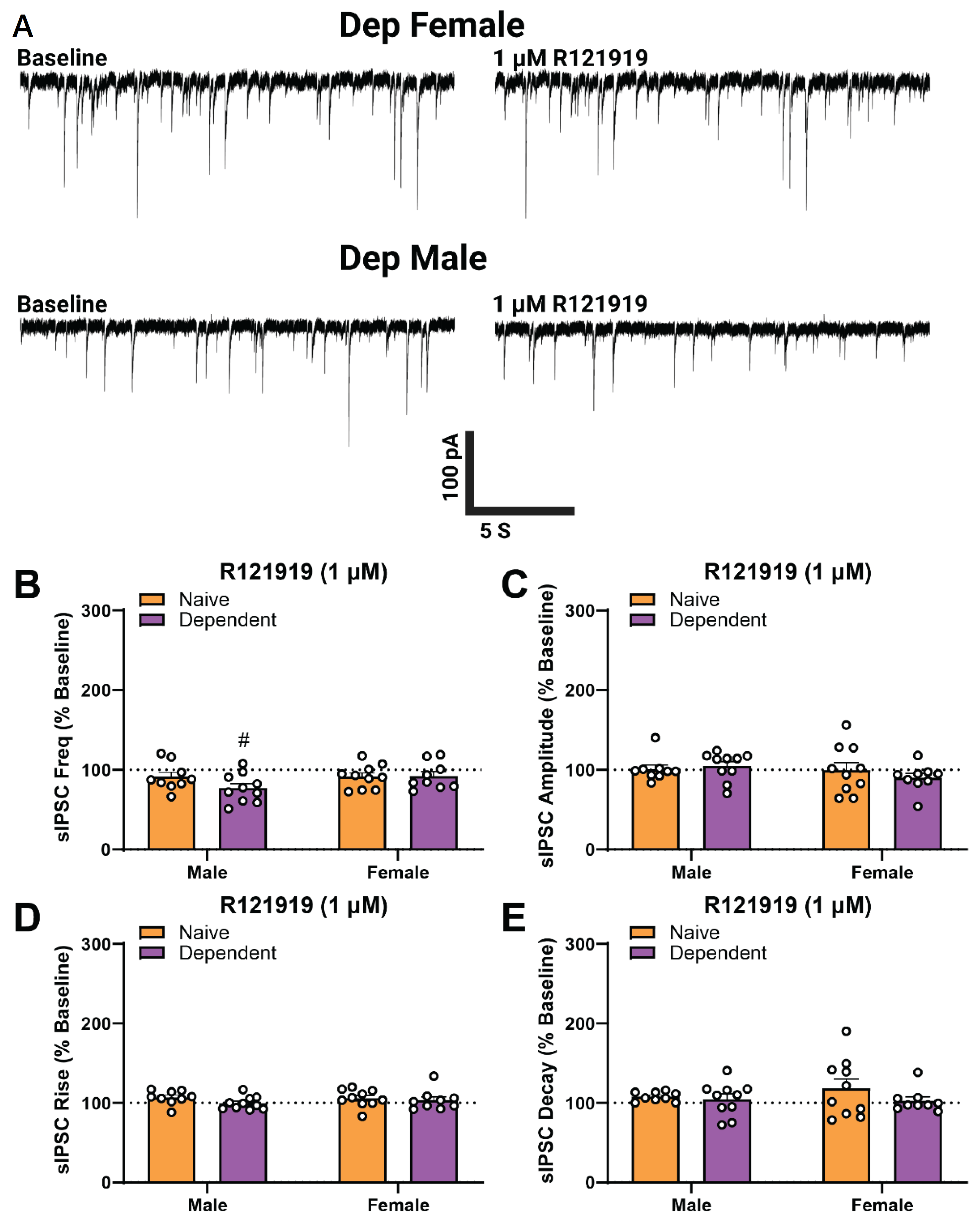
2.4. Alcohol Dependence Induces Tonic Activation of CRF1 at CeA GABAergic Synapses in Males Only
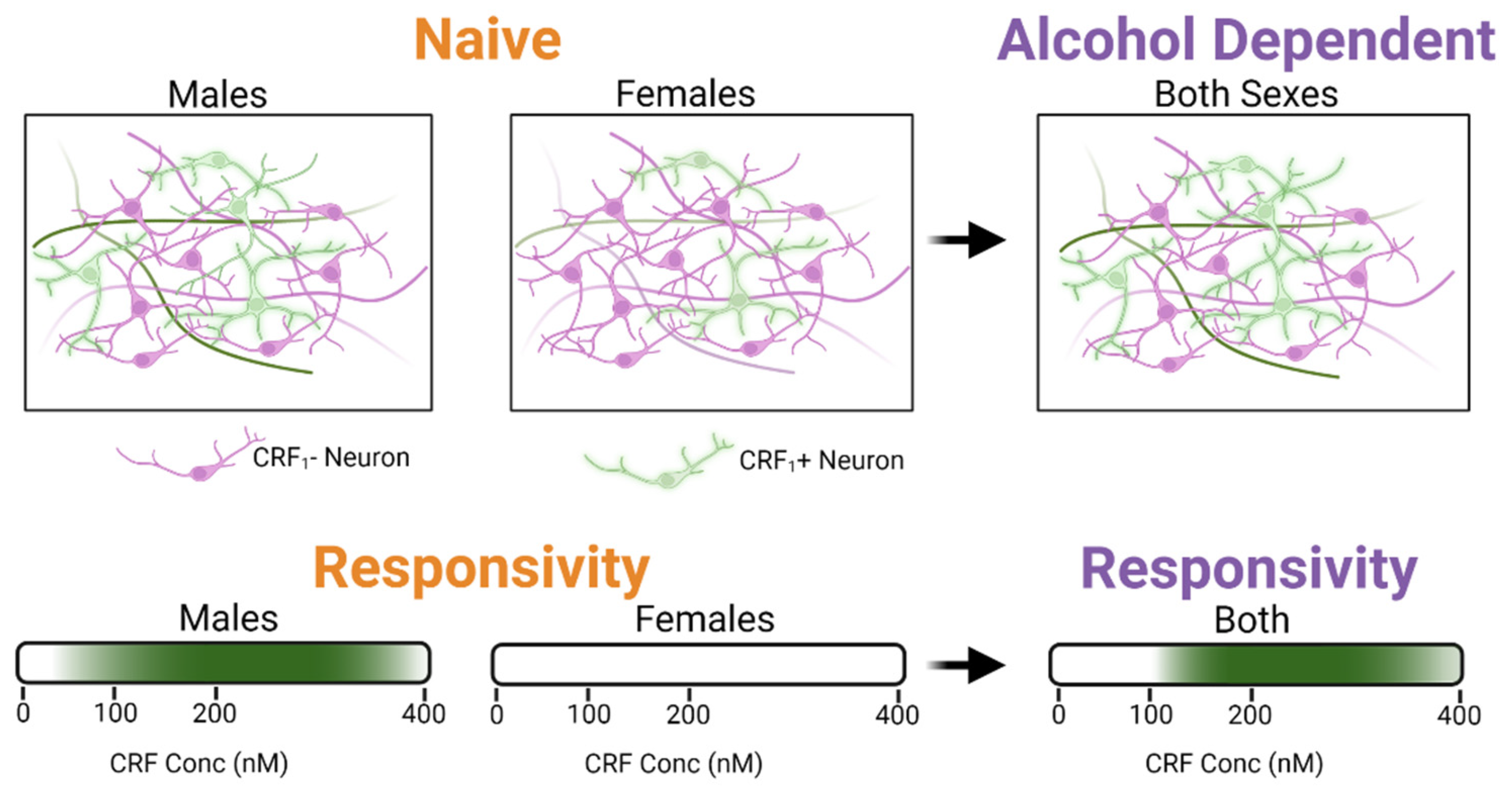
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Electrophysiology
4.3. Drugs
4.4. In Situ Hybridization and Confocal Microscopy
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alcohol Exposure | Sex | sIPSC Property | Baseline | 100 nM CRF | Baseline | 200 nM CRF | Baseline | 400 nM CRF | Baseline | R121919 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |||
| Naïve | Male | Frequency (Hz) | 2.99 | 0.68 | 3.93 | 0.99 | 1.13 | 0.23 | 1.85 | 0.38 | 3.14 | 0.90 | 3.63 | 1.02 | 3.16 | 0.77 | 2.86 | 0.68 |
| Amplitude (pA) | 62.98 | 7.63 | 61.77 | 6.76 | 70.07 | 10.07 | 66.88 | 6.86 | 59.91 | 6.25 | 56.49 | 5.68 | 79.49 | 9.00 | 81.41 | 10.59 | ||
| Rise Time (ms) | 2.57 | 0.13 | 2.59 | 0.11 | 2.61 | 0.14 | 2.54 | 0.10 | 2.55 | 0.08 | 2.61 | 0.12 | 2.33 | 0.08 | 2.49 | 0.11 | ||
| Decay Time (ms) | 9.46 | 1.16 | 9.68 | 0.84 | 9.68 | 0.84 | 8.23 | 1.04 | 9.41 | 0.85 | 9.61 | 0.80 | 10.28 | 1.27 | 11.32 | 1.43 | ||
| n | 9 | 12 | 11 | 9 | ||||||||||||||
| Female | Frequency (Hz) | 1.48 | 0.36 | 1.52 | 0.28 | 1.73 | 0.58 | 1.61 | 0.49 | 1.52 | 0.34 | 1.94 | 0.58 | 2.25 | 0.58 | 2.01 | 0.51 | |
| Amplitude (pA) | 66.42 | 5.97 | 64.47 | 6.52 | 65.71 | 5.13 | 66.58 | 4.94 | 57.95 | 5.77 | 69.99 | 9.01 | 68.21 | 6.96 | 65.28 | 6.99 | ||
| Rise Time (ms) | 2.73 | 0.11 | 2.60 | 0.11 | 2.04 | 0.11 | 2.19 | 0.15 | 2.65 | 0.12 | 2.65 | 0.10 | 2.84 | 0.16 | 3.01 | 0.19 | ||
| Decay Time (ms) | 7.47 | 0.82 | 7.77 | 0.87 | 5.92 | 0.81 | 6.57 | 1.03 | 7.76 | 1.27 | 8.74 | 1.19 | 9.95 | 0.98 | 11.30 | 1.07 | ||
| n | 10 | 9 | 8 | 10 | ||||||||||||||
| Dependent | Male | Frequency (Hz) | 3.30 | 0.40 | 3.56 | 0.47 | 1.43 | 0.29 | 2.25 | 0.68 | 2.63 | 0.54 | 3.14 | 0.68 | 1.79 | 0.42 | 1.48 | 0.37 |
| Amplitude (pA) | 85.01 | 10.06 | 83.44 | 13.25 | 61.75 | 7.82 | 62.05 | 7.78 | 80.04 | 10.23 | 78.06 | 12.82 | 57.55 | 6.56 | 59.45 | 6.47 | ||
| Rise Time (ms) | 2.42 | 0.06 | 2.48 | 0.06 | 2.63 | 0.09 | 2.61 | 0.10 | 2.56 | 0.07 | 2.56 | 0.06 | 3.14 | 0.16 | 3.09 | 0.18 | ||
| Decay Time (ms) | 10.20 | 1.03 | 10.16 | 0.92 | 9.71 | 1.44 | 8.87 | 1.28 | 10.61 | 0.72 | 9.91 | 0.87 | 12.68 | 1.20 | 13.04 | 1.02 | ||
| n | 9 | 9 | 11 | 10 | ||||||||||||||
| Female | Frequency (Hz) | 2.42 | 0.56 | 2.42 | 0.56 | 1.50 | 0.21 | 2.57 | 0.57 | 1.63 | 0.23 | 2.13 | 0.37 | 1.93 | 0.62 | 1.68 | 0.48 | |
| Amplitude (pA) | 59.18 | 3.40 | 53.49 | 2.23 | 95.01 | 15.07 | 112.62 | 21.57 | 62.84 | 8.16 | 68.50 | 15.50 | 71.30 | 12.03 | 62.58 | 9.93 | ||
| Rise Time (ms) | 2.82 | 0.25 | 2.73 | 0.27 | 2.23 | 0.09 | 2.29 | 0.07 | 2.61 | 0.19 | 2.72 | 0.20 | 3.10 | 0.18 | 3.19 | 0.16 | ||
| Decay Time (ms) | 12.09 | 1.15 | 10.84 | 1.17 | 8.45 | 0.88 | 9.15 | 1.11 | 10.81 | 0.86 | 10.14 | 0.95 | 11.47 | 1.28 | 11.70 | 1.14 | ||
| n | 8 | 11 | 9 | 9 | ||||||||||||||
References
- Koob, G.F.; Buck, C.L.; Cohen, A.; Edwards, S.; Park, P.E.; Schlosburg, J.E.; Schmeichel, B.; Vendruscolo, L.F.; Wade, C.L.; Whitfield, T.W., Jr.; et al. Addiction as a stress surfeit disorder. Neuropharmacology 2014, 76 Pt B, 370–382. [Google Scholar] [CrossRef] [Green Version]
- Koob, G.F. Anhedonia, Hyperkatifeia, and Negative Reinforcement in Substance Use Disorders. In Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Zorrilla, E.P.; Koob, G.F. Progress in corticotropin-releasing factor-1 antagonist development. Drug Discov. Today 2010, 15, 371–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilpin, N.W.; Roberto, M. Neuropeptide modulation of central amygdala neuroplasticity is a key mediator of alcohol dependence. Neurosci. Biobehav. Rev. 2012, 36, 873–888. [Google Scholar] [CrossRef] [Green Version]
- Lowery, E.G.; Thiele, T.E. Pre-clinical evidence that corticotropin-releasing factor (CRF) receptor antagonists are promising targets for pharmacological treatment of alcoholism. CNS Neurol. Disord. Drug Targets 2010, 9, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Roberto, M.; Spierling, S.R.; Kirson, D.; Zorrilla, E.P. Corticotropin-Releasing Factor (CRF) and Addictive Behaviors. Int. Rev. Neurobiol. 2017, 136, 5–51. [Google Scholar] [CrossRef] [PubMed]
- Quadros, I.M.; Macedo, G.C.; Domingues, L.P.; Favoretto, C.A. An Update on CRF Mechanisms Underlying Alcohol Use Disorders and Dependence. Front. Endocrinol. 2016, 7, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberto, M.; Gilpin, N.W.; Siggins, G.R. The central amygdala and alcohol: Role of Gamma-Aminobutyric acid, glutamate, and neuropeptides. Cold Spring Harb. Perspect. Med. 2012, 2, a012195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koob, G.F. Theoretical frameworks and mechanistic aspects of alcohol addiction: Alcohol addiction as a reward deficit disorder. Curr. Top. Behav. Neurosci. 2013, 13, 3–30. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, V.A.; Herman, M.A.; Cuzon Carlson, V.C.; Walter, N.A.; Grant, K.A.; Roberto, M. Synaptic adaptations in the central amygdala and hypothalamic paraventricular nucleus associated with protracted ethanol abstinence in male rhesus monkeys. Neuropsychopharmacology 2019, 44, 982–993. [Google Scholar] [CrossRef]
- Roberto, M.; Kirson, D.; Khom, S. The Role of the Central Amygdala in Alcohol Dependence. Cold Spring Harb. Perspect. Med. 2021, 11, a039339. [Google Scholar] [CrossRef] [Green Version]
- Bale, T.L.; Vale, W.W. CRF and CRF receptors: Role in stress responsivity and other behaviors. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 525–557. [Google Scholar] [CrossRef] [PubMed]
- Hauger, R.L.; Risbrough, V.; Brauns, O.; Dautzenberg, F.M. Corticotropin releasing factor (CRF) receptor signaling in the central nervous system: New molecular targets. CNS Neurol. Disord. Drug Targets 2006, 5, 453–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilig, M.; Koob, G.F. A key role for corticotropin-releasing factor in alcohol dependence. Trends Neurosci. 2007, 30, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Fekete, E.M.; Zorrilla, E.P. Physiology, pharmacology, and therapeutic relevance of urocortins in mammals: Ancient CRF paralogs. Front. Neuroendocrinol. 2007, 28, 1–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlo Pich, E.; Lorang, M.; Yeganeh, M.; Rodriguez de Fonseca, F.; Raber, J.; Koob, G.F.; Weiss, F. Increase of extracellular corticotropin-releasing factor-like immunoreactivity levels in the amygdala of awake rats during restraint stress and ethanol withdrawal as measured by microdialysis. J. Neurosci. 1995, 15, 5439–5447. [Google Scholar] [CrossRef] [Green Version]
- Roberto, M.; Cruz, M.T.; Gilpin, N.W.; Sabino, V.; Schweitzer, P.; Bajo, M.; Cottone, P.; Madamba, S.G.; Stouffer, D.G.; Zorrilla, E.P.; et al. Corticotropin releasing factor-induced amygdala Gamma-Aminobutyric Acid release plays a key role in alcohol dependence. Biol. Psychiatry 2010, 67, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Valdez, G.R.; Zorrilla, E.P.; Roberts, A.J.; Koob, G.F. Antagonism of corticotropin-releasing factor attenuates the enhanced responsiveness to stress observed during protracted ethanol abstinence. Alcohol 2003, 29, 55–60. [Google Scholar] [CrossRef]
- Gehlert, D.R.; Cippitelli, A.; Thorsell, A.; Le, A.D.; Hipskind, P.A.; Hamdouchi, C.; Lu, J.; Hembre, E.J.; Cramer, J.; Song, M.; et al. 3-(4-Chloro-2-morpholin-4-yl-thiazol-5-yl)-8-(1-ethylpropyl)-2,6-dimethyl-imidazo [1,2-b]pyridazine: A novel brain-penetrant, orally available corticotropin-releasing factor receptor 1 antagonist with efficacy in animal models of alcoholism. J. Neurosci. 2007, 27, 2718–2726. [Google Scholar] [CrossRef]
- Richardson, H.N.; Zhao, Y.; Fekete, E.M.; Funk, C.K.; Wirsching, P.; Janda, K.D.; Zorrilla, E.P.; Koob, G.F. MPZP: A novel small molecule corticotropin-releasing factor type 1 receptor (CRF1) antagonist. Pharmacol. Biochem. Behav. 2008, 88, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Overstreet, D.H.; Knapp, D.J.; Breese, G.R. Similar anxiety-like responses in male and female rats exposed to repeated withdrawals from ethanol. Pharmacol. Biochem. Behav. 2004, 78, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Kwako, L.E.; Spagnolo, P.A.; Schwandt, M.L.; Thorsell, A.; George, D.T.; Momenan, R.; Rio, D.E.; Huestis, M.; Anizan, S.; Concheiro, M.; et al. The corticotropin releasing hormone-1 (CRH1) receptor antagonist pexacerfont in alcohol dependence: A randomized controlled experimental medicine study. Neuropsychopharmacology 2015, 40, 1053–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwandt, M.L.; Cortes, C.R.; Kwako, L.E.; George, D.T.; Momenan, R.; Sinha, R.; Grigoriadis, D.E.; Pich, E.M.; Leggio, L.; Heilig, M. The CRF1 Antagonist Verucerfont in Anxious Alcohol-Dependent Women: Translation of Neuroendocrine, But not of Anti-Craving Effects. Neuropsychopharmacology 2016, 41, 2818–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, M.Q.; Kirson, D.; Wolfe, S.A.; Khom, S.; D’Ambrosio, S.R.; Spierling Bagsic, S.R.; Bajo, M.; Vlkolinsky, R.; Hoang, N.K.; Singhal, A.; et al. Importance of sex and trauma context on circulating cytokines and amygdalar GABAergic signaling in a comorbid model of posttraumatic stress and alcohol use disorders. Mol. Psychiatry 2021, 26, 3093–3107. [Google Scholar] [CrossRef] [PubMed]
- Kirson, D.; Steinman, M.Q.; Wolfe, S.A.; Spierling Bagsic, S.R.; Bajo, M.; Sureshchandra, S.; Oleata, C.S.; Messaoudi, I.; Zorrilla, E.P.; Roberto, M. Sex and context differences in the effects of trauma on comorbid alcohol use and post-traumatic stress phenotypes in actively drinking rats. J. Neurosci. Res. 2021, 99, 3354–3372. [Google Scholar] [CrossRef]
- Shansky, R.M. Sex differences in PTSD resilience and susceptibility: Challenges for animal models of fear learning. Neurobiol. Stress 2015, 1, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Wellman, C.L.; Bangasser, D.A.; Bollinger, J.L.; Coutellier, L.; Logrip, M.L.; Moench, K.M.; Urban, K.R. Sex Differences in Risk and Resilience: Stress Effects on the Neural Substrates of Emotion and Motivation. J. Neurosci. 2018, 38, 9423–9432. [Google Scholar] [CrossRef] [Green Version]
- Cover, K.K.; Maeng, L.Y.; Lebron-Milad, K.; Milad, M.R. Mechanisms of estradiol in fear circuitry: Implications for sex differences in psychopathology. Transl. Psychiatry 2014, 4, e422. [Google Scholar] [CrossRef] [Green Version]
- Agoglia, A.E.; Tella, J.; Herman, M.A. Sex differences in corticotropin releasing factor peptide regulation of inhibitory control and excitability in central amygdala corticotropin releasing factor receptor 1-neurons. Neuropharmacology 2020, 180, 108296. [Google Scholar] [CrossRef]
- Kirson, D.; Khom, S.; Rodriguez, L.; Wolfe, S.A.; Varodayan, F.P.; Gandhi, P.J.; Patel, R.R.; Vlkolinsky, R.; Bajo, M.; Roberto, M. Sex Differences in Acute Alcohol Sensitivity of Naive and Alcohol Dependent Central Amygdala GABA Synapses. Alcohol Alcohol. 2021, 56, 581–588. [Google Scholar] [CrossRef]
- Nie, Z.; Schweitzer, P.; Roberts, A.J.; Madamba, S.G.; Moore, S.D.; Siggins, G.R. Ethanol augments GABAergic transmission in the central amygdala via CRF1 receptors. Science 2004, 303, 1512–1514. [Google Scholar] [CrossRef]
- Nie, Z.; Zorrilla, E.P.; Madamba, S.G.; Rice, K.C.; Roberto, M.; Siggins, G.R. Presynaptic CRF1 receptors mediate the ethanol enhancement of GABAergic transmission in the mouse central amygdala. Sci. World J. 2009, 9, 68–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varodayan, F.P.; Correia, D.; Kirson, D.; Khom, S.; Oleata, C.S.; Luu, G.; Schweitzer, P.; Roberto, M. CRF modulates glutamate transmission in the central amygdala of naive and ethanol-dependent rats. Neuropharmacology 2017, 125, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Varodayan, F.P.; Logrip, M.L.; Roberto, M. P/Q-type voltage-gated calcium channels mediate the ethanol and CRF sensitivity of central amygdala GABAergic synapses. Neuropharmacology 2017, 125, 197–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varodayan, F.P.; de Guglielmo, G.; Logrip, M.L.; George, O.; Roberto, M. Alcohol Dependence Disrupts Amygdalar L-Type Voltage-Gated Calcium Channel Mechanisms. J. Neurosci. 2017, 37, 4593–4603. [Google Scholar] [CrossRef] [Green Version]
- Rouzer, S.K.; Diaz, M.R. Factors of sex and age dictate the regulation of GABAergic activity by corticotropin-releasing factor receptor 1 in the medial sub-nucleus of the central amygdala. Neuropharmacology 2021, 189, 108530. [Google Scholar] [CrossRef]
- Agoglia, A.E.; Zhu, M.; Quadir, S.G.; Bluitt, M.N.; Douglass, E.; Hanback, T.; Tella, J.; Ying, R.; Hodge, C.W.; Herman, M.A. Sex-specific plasticity in CRF regulation of inhibitory control in central amygdala CRF1 neurons after chronic voluntary alcohol drinking. Addict. Biol. 2022, 27, e13067. [Google Scholar] [CrossRef]
- Retson, T.A.; Hoek, J.B.; Sterling, R.C.; Van Bockstaele, E.J. Amygdalar neuronal plasticity and the interactions of alcohol, sex, and stress. Brain Struct. Funct. 2015, 220, 3211–3232. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Roberto, M. Cell-type-specific tonic GABA signaling in the rat central amygdala is selectively altered by acute and chronic ethanol. Addict. Biol. 2016, 21, 72–86. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.A.; Contet, C.; Roberto, M. A Functional Switch in Tonic GABA Currents Alters the Output of Central Amygdala Corticotropin Releasing Factor Receptor-1 Neurons Following Chronic Ethanol Exposure. J. Neurosci. 2016, 36, 10729–10741. [Google Scholar] [CrossRef]
- Augier, E.; Barbier, E.; Dulman, R.S.; Licheri, V.; Augier, G.; Domi, E.; Barchiesi, R.; Farris, S.; Natt, D.; Mayfield, R.D.; et al. A molecular mechanism for choosing alcohol over an alternative reward. Science 2018, 360, 1321–1326. [Google Scholar] [CrossRef] [Green Version]
- Kreifeldt, M.; Herman, M.A.; Sidhu, H.; Okhuarobo, A.; Macedo, G.C.; Shahryari, R.; Gandhi, P.J.; Roberto, M.; Contet, C. Central amygdala corticotropin-releasing factor neurons promote hyponeophagia but do not control alcohol drinking in mice. Mol. Psychiatry 2022, 27, 2502–2513. [Google Scholar] [CrossRef]
- Patel, R.R.; Varodayan, F.P.; Herman, M.A.; Jimenez, V.; Agnore, R.; Gao, L.; Bajo, M.; Cuzon Carlson, V.C.; Walter, N.A.; Fei, S.S.; et al. Synaptic effects of IL-1beta and CRF in the central amygdala after protracted alcohol abstinence in male rhesus macaques. Neuropsychopharmacology 2022, 47, 847–856. [Google Scholar] [CrossRef]
- Pomrenze, M.B.; Millan, E.Z.; Hopf, F.W.; Keiflin, R.; Maiya, R.; Blasio, A.; Dadgar, J.; Kharazia, V.; De Guglielmo, G.; Crawford, E.; et al. A Transgenic Rat for Investigating the Anatomy and Function of Corticotrophin Releasing Factor Circuits. Front. Neurosci. 2015, 9, 487. [Google Scholar] [CrossRef] [PubMed]
- Pomrenze, M.B.; Giovanetti, S.M.; Maiya, R.; Gordon, A.G.; Kreeger, L.J.; Messing, R.O. Dissecting the Roles of GABA and Neuropeptides from Rat Central Amygdala CRF Neurons in Anxiety and Fear Learning. Cell Rep. 2019, 29, 13–21.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Guglielmo, G.; Kallupi, M.; Pomrenze, M.B.; Crawford, E.; Simpson, S.; Schweitzer, P.; Koob, G.F.; Messing, R.O.; George, O. Inactivation of a CRF-dependent amygdalofugal pathway reverses addiction-like behaviors in alcohol-dependent rats. Nat. Commun. 2019, 10, 1238. [Google Scholar] [CrossRef] [PubMed]
- Pleil, K.E.; Rinker, J.A.; Lowery-Gionta, E.G.; Mazzone, C.M.; McCall, N.M.; Kendra, A.M.; Olson, D.P.; Lowell, B.B.; Grant, K.A.; Thiele, T.E.; et al. NPY signaling inhibits extended amygdala CRF neurons to suppress binge alcohol drinking. Nat. Neurosci. 2015, 18, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Kirson, D.; Oleata, C.S.; Roberto, M. Taurine Suppression of Central Amygdala GABAergic Inhibitory Signaling via Glycine Receptors Is Disrupted in Alcohol Dependence. Alcohol. Clin. Exp. Res. 2020, 44, 445–454. [Google Scholar] [CrossRef]
- Khom, S.; Nguyen, J.D.; Vandewater, S.A.; Grant, Y.; Roberto, M.; Taffe, M.A. Self-Administration of Entactogen Psychostimulants Dysregulates Gamma-Aminobutyric Acid (GABA) and Kappa Opioid Receptor Signaling in the Central Nucleus of the Amygdala of Female Wistar Rats. Front. Behav. Neurosci. 2021, 15, 780500. [Google Scholar] [CrossRef]
- Khom, S.; Steinkellner, T.; Hnasko, T.S.; Roberto, M. Alcohol dependence potentiates substance P/neurokinin-1 receptor signaling in the rat central nucleus of amygdala. Sci. Adv. 2020, 6, eaaz1050. [Google Scholar] [CrossRef]
- Khom, S.; Rodriguez, L.; Gandhi, P.; Kirson, D.; Bajo, M.; Oleata, C.S.; Vendruscolo, L.F.; Mason, B.J.; Roberto, M. Alcohol dependence and withdrawal increase sensitivity of central amygdalar GABAergic synapses to the glucocorticoid receptor antagonist mifepristone in male rats. Neurobiol. Dis. 2022, 164, 105610. [Google Scholar] [CrossRef]
- Varodayan, F.P.; Patel, R.R.; Matzeu, A.; Wolfe, S.A.; Curley, D.E.; Khom, S.; Gandhi, P.J.; Rodriguez, L.; Bajo, M.; D’Ambrosio, S.; et al. The Amygdala Noradrenergic System Is Compromised with Alcohol Use Disorder. Biol. Psychiatry 2022, 91, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, S.A.; Sidhu, H.; Patel, R.R.; Kreifeldt, M.; D’Ambrosio, S.R.; Contet, C.; Roberto, M. Molecular, Morphological, and Functional Characterization of Corticotropin-Releasing Factor Receptor 1-Expressing Neurons in the Central Nucleus of the Amygdala. eNeuro 2019, 6, e0087-19.2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, T.R.; Kang, I.H.; Wheeler, D.B.; Lindquist, R.A.; Papallo, A.; Sabatini, D.M.; Golland, P.; Carpenter, A.E. CellProfiler Analyst: Data exploration and analysis software for complex image-based screens. BMC Bioinform. 2008, 9, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, L.; Kirson, D.; Wolfe, S.A.; Patel, R.R.; Varodayan, F.P.; Snyder, A.E.; Gandhi, P.J.; Khom, S.; Vlkolinsky, R.; Bajo, M.; et al. Alcohol Dependence Induces CRF Sensitivity in Female Central Amygdala GABA Synapses. Int. J. Mol. Sci. 2022, 23, 7842. https://doi.org/10.3390/ijms23147842
Rodriguez L, Kirson D, Wolfe SA, Patel RR, Varodayan FP, Snyder AE, Gandhi PJ, Khom S, Vlkolinsky R, Bajo M, et al. Alcohol Dependence Induces CRF Sensitivity in Female Central Amygdala GABA Synapses. International Journal of Molecular Sciences. 2022; 23(14):7842. https://doi.org/10.3390/ijms23147842
Chicago/Turabian StyleRodriguez, Larry, Dean Kirson, Sarah A. Wolfe, Reesha R. Patel, Florence P. Varodayan, Angela E. Snyder, Pauravi J. Gandhi, Sophia Khom, Roman Vlkolinsky, Michal Bajo, and et al. 2022. "Alcohol Dependence Induces CRF Sensitivity in Female Central Amygdala GABA Synapses" International Journal of Molecular Sciences 23, no. 14: 7842. https://doi.org/10.3390/ijms23147842