Competitive games among plants
 It has long been recognized that individual-level selection can lead to sub-optimal outcomes when individuals have shared access to resources. In the classic example of the tragedy of the commons (described in Hardin 1968) herders will tend to seriously overgraze shared land. This occurs because each individual herder, with the ability to add another sheep to the field, will get the benefits of that additional sheep while the costs of the additional overgrazing will be shared among the group.
It has long been recognized that individual-level selection can lead to sub-optimal outcomes when individuals have shared access to resources. In the classic example of the tragedy of the commons (described in Hardin 1968) herders will tend to seriously overgraze shared land. This occurs because each individual herder, with the ability to add another sheep to the field, will get the benefits of that additional sheep while the costs of the additional overgrazing will be shared among the group.
Tragedies of the commons occur in plant communities as well. Plants have shared access to light, water and nutrients. When one tree grows tall or invests a lot in fine roots, its neighbors must step up and do the same in order to gain equal access to light, water and nutrients. Investments like these, which are necessary for an individuals’ competitive ability but that can harm the productivity of the group can be called “competitive overinvestments”.
Evidence of competitive overinvestments has been demonstrated directly. Gersani et al. (2001) put together a soybean experiment where they grew beans in either shared pots of separate pots with the same amount of resource/individual in each. In the shared pots, where plants were under the influence of competition, individuals invested more in fine roots at the cost of investment in seeds and thus reproduction. We have also seen evidence in competitive overinvestments indirectly in qualitative model data comparisons to explain: major tradeoff axes in allocation strategies across forests (Farrior et al. 2013a) and in complex plant responses to nitrogen and water addition experiments (Farrior et al. 2013b).
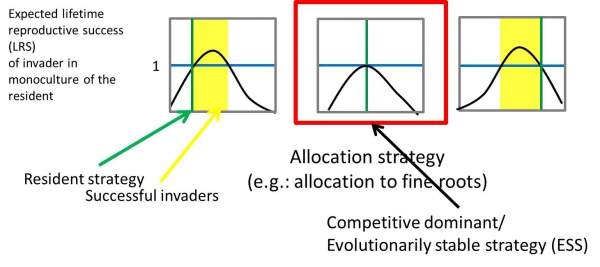
Below is a simple example from Farrior (2014) meant to demonstrate the influence of sharing access to resources on dominant plant strategies and the resulting decline in community resource use efficiency.
| Simple example of competitive overinvestments in plants |
|
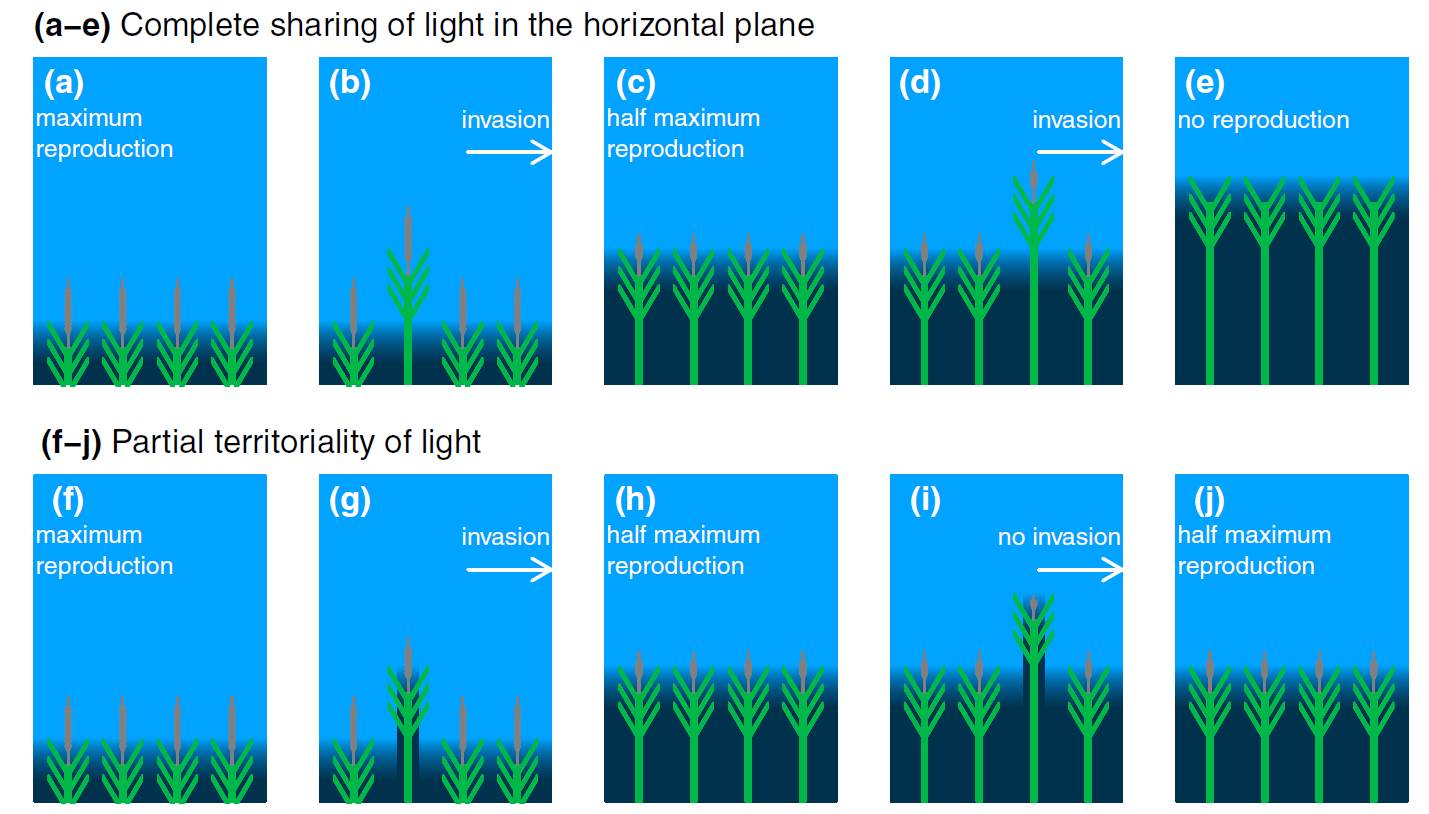
If resources are shared, optimization of individual-level fitness often leads to sub-optimal community resource-use efficiency, a tragedy of the commons (Hardin, 1968). Here are two examples of hypothetical processes of evolution (or community assembly) for plants that share light completely (a–e) and for plants that only share half of their light (f–j) in the horizontal plane. Individual plants are depicted as a green stem and leaves. Light level is indicated by the brightness of the blue background. Individuals differ in investment in height. To indicate fitness, reproductive output is depicted as seed head length and is proportional to light uptake minus height. Note these examples assume that each community (panel) is large enough that any one individual does not have a measurable effect on the resource level of other individuals. The end point of evolution (or community assembly) for each scenario is pictured in (e) and (j). For the complete-sharing individuals, a dead-end community with
|
 |
|
no reproduction (e) is arrived at via successive invasions of individuals with greater investment in height (b and d). Increases in height pay off to individuals in competition with shorter individuals, but lead to diminished fitness as the taller individuals come to dominate the community. In competition with one another, these individuals take up the same amount of light as their shorter counterparts, but have greater height costs (compare a, c, and e). If resource uptake is partially territorial however, the end point of evolution is a viable community (j). Here, individuals that are taller than their neighbors (in g and i) receive only half the benefits as they had when resources are shared completely (in b and d) because of the presence of their own self shading. This leads to a community in (h) that cannot be invaded by individuals of greater height (j = h; the taller invader in (i) is unsuccessful). Here self-shading caused by partial territoriality restrains the community from the dead-end scenario in the upper panels. Yet, because resource sharing is still present, the stable community still only has a fraction of the maximum community level resource-use efficiency and reproduction (a and f). Farrior 2014, New Phytologist 203:1025-1027, Fig. 1. |