by Josef Spacek
Ribosomes are spheroid or ellipsoid particles about 15 x 25 nm in diameter, consisting of rRNA and protein. They are formed by two subunits. Single ribosomes are inactive. When linked into polyribosomes (polysomes), they read and translate from mRNA a message transcripted from nuclear DNA and synthesize peptides or proteins from amino acids according to this genetic instruction (Fig. 1). In electron micrographs, ribosomes should be strictly differentiated from glycogen beta-particles of nearly the same diameter (15 - 30 nm) (Fig. 2).
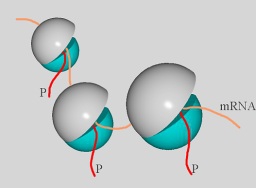
Fig. 1: Ribosomes synthesize proteins (red) according to instruction translated from mRNA (yellow).
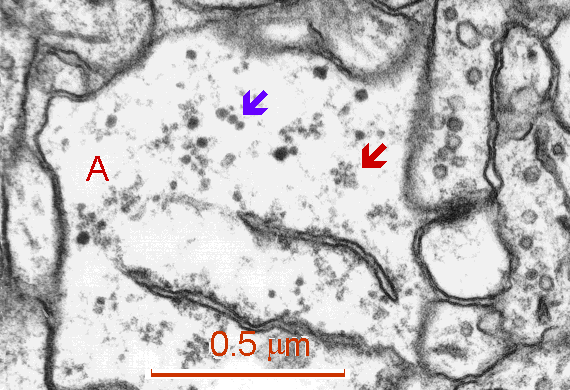
Fig. 2: Free polyribosomes (red arrow) and glycogen beta-particles (blue arrow) in a process of astrocyte (A). Cisterns of granular endoplasmic reticulum are present in the lower part of figure. (Rat, hippocampus, CA1)
Polyribosomes either lie free, releasing new peptides into the cytoplasm, or are attached to the membrane of endoplasmic reticulum (rough, granular e.r.), releasing new peptides into its cisternal or tubular lumen (Weiss 1988).
The synapse is constructed from receptors, ion channels, adhesion molecules, second messenger systems (e.g., protein kinase C which triggers a variety of protein phosphorylations, protein kinase II which participates in Ca2+ signaling in tandem with calmodulin and inositol 1,4,5-trisphophate which mobilizes Ca2+ from stores such as smooth endoplasmic reticulum), and cytoskeletal and associated proteins (such as actin, myosin, tubulin, MAP2, etc.). Most of these components are highly specialized proteins. They are not transported into thousands of synaptic sites from the neuronal soma but particular mRNA molecules are differentially distributed via selective intracellular transport from the nucleus to the postsynaptic microdomains. Here they form the machinery which allows a local synthesis of specific synaptic proteins (Steward and Banker 1992). Subsynaptic regions are thus typical representatives of proteosynthesis independent of neuronal soma.
Polyribosomes are present beneath the various types of synapses on dendrite shafts (symmetric, asymmetric - Fig. 3 a,b,c), axonal initial segments (inhibitory), and especially beneath the synapses present on dendritic spines. They are found in spine heads (Fig. 4 a,b) and necks ( Fig. 5), but most of them (80%) form clusters beneath spines, in the "mounds" of the spine bases (Figs. 6 a,b, 7).
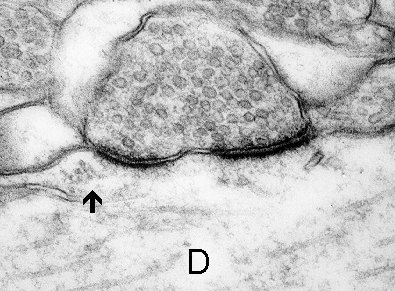
3a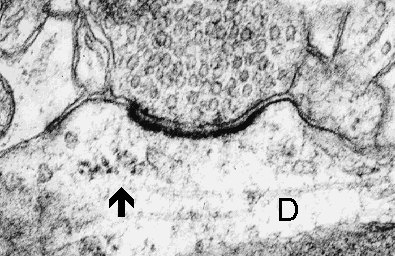
3b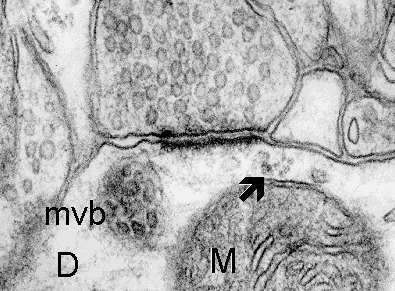
3c
Fig. 3 a,b,c: A cluster of polyribosomes (arrow) in the subsynaptic region of synapses formed on the dendrite (D). Mitochondrion (M) and multivesicular body (mvb) are associated with the subsynaptic region in c. (Rat, hippocampus, CA1)
4a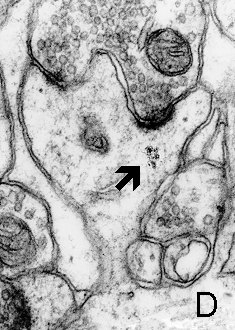
4b
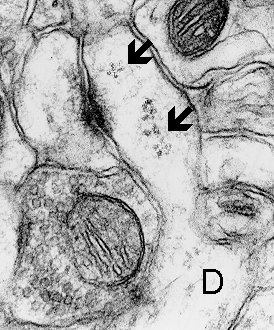
Fig. 4 a,b: Polyribosomes (arrows) in the heads of dendritic spines. a. Thin spine (mouse, neocortex) b. Mushroom spine (rat, hippocampus, CA1). D - dendrite.
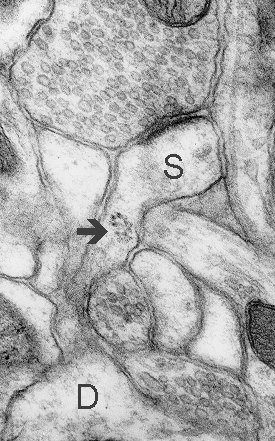
Fig. 5: A cluster of polyribosomes (arrow) in a neck of a thin spine (S). D - dendrite. (Rat, hippocampus, CA1)
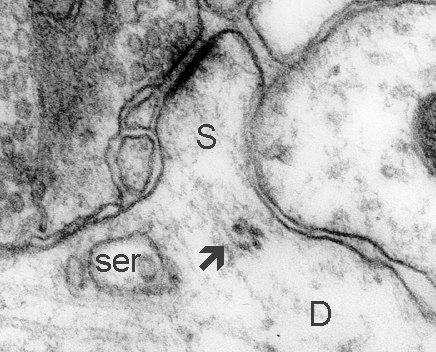
Fig. 6: Polyribosomes (arrow) at the base of a short, sessile spine. D - dendrite, ser - smooth endoplasmic reticulum. (Rat, hippocampus, CA1)
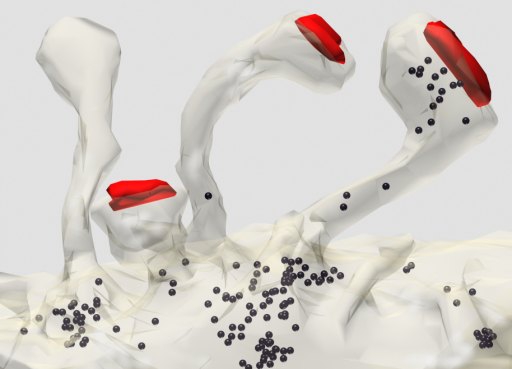
Fig. 7: Three-dimensional reconstruction of thin spines emerging from a dendrite (made transparent). Polyribosomes (black) are most frequent at the bases of spines. Areas of synaptic contact are shown in red. (Rat, hippocampus, CA1)
50% of the subsynaptic polyribosomes are found in association with tubules of endoplasmic reticulum (Steward and Reeves 1988) (Fig. 8), including the spine apparatus deriving from it (Fig. 9 a,b,c). The term "synapse-associated polyribosomal complex" has been proposed for this association (Steward, Falk and Torre 1996). It is hypothesized that the reticulum may be involved in post-translational processing of proteins synthesized by the postsynaptic polyribosomes.
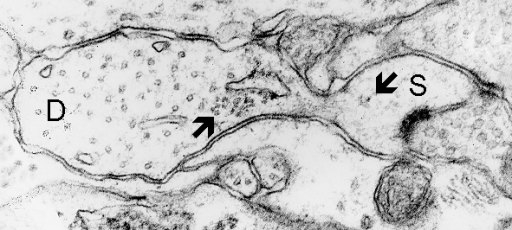
Fig. 8: Polyribosomes both free in the head of spine (S) and associated with endoplasmic reticulum (arrows) at the base of a dendritic spine. D - dendrite. (Rat, hippocampus, CA1)
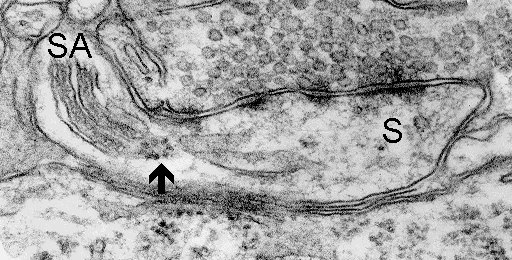
9a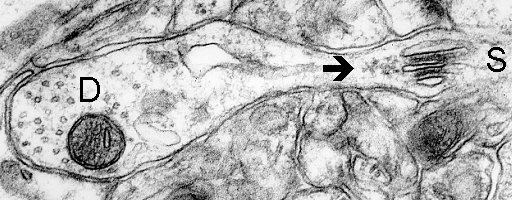
9b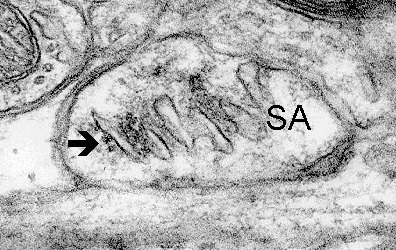
9c
Fig. 9 a,b,c: Polyribosomes (arrows) closely associated with spine apparatuses (SA). D - dendrite, S - spine. (Mouse, neocortex)
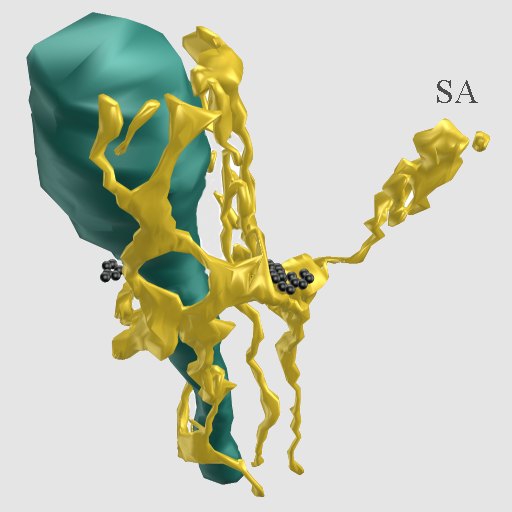
Fig. 10: Three-dimensional reconstruction of endoplasmic reticulum (yellow) of a dendrite and spine. The spine apparatus (SA) is connected by a narrow tubule with a cistern at the base of the spine, associated with this is a cluster of polyribosomes (black). A nearby mitochondrion is colored dark green. (Rat, hippocampus, CA1)
A dramatic increase in polyribosomes associated with dendritic spines was observed during periods of post-lesion synaptogenesis induced by reinnervation in denervated dendrites. The number of polyribosomes in spine heads, necks, and at their bases increased about 3-fold, presumably to produce proteins involved in synapse growth (Steward 1983; Steward and Fass 1983; Steward and Falk, 1985). Enormous numbers of polyribosomal clusters were found in the bases of complex branched spines of thalamocortical relay neurons (Spacek and Lieberman 1974) (Fig. 11 a,b).
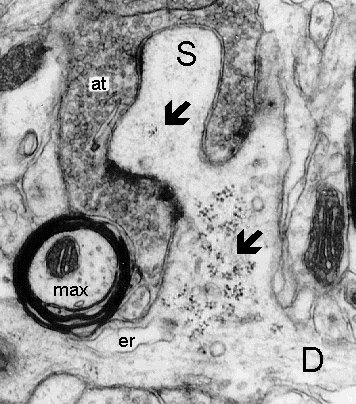
11a
11b
Fig. 11: a. A widened base of a multisynaptic excrescence (S) contains an enormous cluster of polyribosomes (lower arrow). b. Three-dimensional reconstruction of the dendrite and excrescence showing the enormous cluster of polyribosomes (black) within. The multiple areas of synaptic contact are shown in red. D - dendrite, er - endoplasmic reticulum, max - myelinated axon, at - giant axon terminal. (Rat, ventrobasal thalamic nucleus)
For a detailed tutorial on identifying polyribosomes in EM images, please see Protocol for Ribosome Identification by L. Ostroff (PDF).
References:
- Spacek J, Lieberman AR (1974) Ultrastructure and three-dimensional organization of synaptic glomeruli in rat somatosensory thalamus. J Anat 117 : 487-516.
- Spacek J, Hartmann M (1983) Three-dimensional analysis of dendritic spines. I. Quantitative observations related to dendritic spines and synaptic morphology in cerebral and cerebellar cortices. Anat Embryol 67 : 299-310.
- Steward O (1983) Alterations in polyribosomes associated with dendritic spines during reinnervation of the detate gyrus of the adult rat. J Neurosci 3, 177-188.
- Steward O, Fass B (1983) Polyribosomes associated with dendritic spines in the denervated dentate gyrus: evidence for local regulation of protein synthesis during reinnervation. In: Changeux J-P, Glowinski J, Imbert M, Bloom FE (eds.): Molecular and Cellular Interactions Underlying Higher Brain Functions. Progr Brain Res 58 : 131-136.
- Steward O, Falk PM (1985) Polyribosomes under developing spine synapses: growth specializations of dendrites at sites of synaptogenesis. J Neurosci Res 13 : 75-88.
- Steward O, Reeves TM (1988) Protein-synthetic machinery beneath postsynaptic sites on CNS neurons: association between polyribosomes and other organelles at the synaptic sites. J Neurosci 8 : 176-184.
- Steward O, Banker GA (1992) getting the message from the gene to the synapse: sorting and intracellular transport of RNA in neurons. TINS 15 : 180-186.
- Steward O, Falk PM and Torre ER (1996) Ultrastructural basis for gene expression at the synapse: synapse-associated polyribosome complexes. J Neurocytol 25 : 717-734.
- Weiss L (ed.) Cell and Tissue Biology. A Textbook of Histology. Urban and Schwarzenberg, Baltimore, Munich, 1988.